A little bit of physiology to start off the summer.
We are gearing up to complete the data sets for local beetles this fall. Ultimately, we’ll be comparing metabolism of two Eleodes species, E. obscura and E. longicollis, with those of a few mimics, Philolithus elatus and Stenomorpha marginata. The data from last fall were very interesting, but after drafting the manuscript it was clear that another season of data would help to clarify the results.
{kind=link}
{kind=link}
Because of seasonal availability, comparing Eleodes with their mimics will have to wait until late summer and early fall. Eleodes longicollis is common all summer, but E. obscura becomes much more common in July. Philolithus elatus and S. marginatus will not appear until the end of July at the earliest.
In the meantime, there were a couple of other species available in the lab that will help in understanding the data from Eleodes, Philolithus, and Stenomorpha.

Zophobas morio, also known as the “superworm,” is originally from the American tropics. It is widely cultured as a food for pets and is being considered as a source of protein for humans. A colony was established in the lab in early 2024, so they are available for experiments any time. Being from the neotropics, with consistently warm and relatively wet conditions, they are expected to respond more strongly to temperature changes.

Asbolus verrucosus, called the blue death-feigning beetle because of its tendency to play dead when handled, is a long-lived species from the hot, dry deserts of southwestern North America. We received a cohort from Bugs in Cyberspace in January, and they have spent the past few months adjusting to handling and conditions in the lab. Their adaptations to extreme conditions suggest that they will be tolerant of high temperatures.
Oxygen consumption was measured for the two species from 15°C, at which beetles should be slow and sluggish, to 40°C, just below the lethal temperature for a close relative of A. verrucosus (Cryptoglossa muricata; Ahearn, 1970). Therefore the temperature range may cause cold stress at at the low end and heat stress on the high end, but was not expected to be lethal.
The two species are just about the same size and mass, with each being somewhat over half a gram. Twelve beetles of each species were tested at each temperature, although technical issues reduced this to eleven in a few cases. The respirometer could handle eight beetles at a time, so four of each species were tested together each day, and each temperature required three days of experiments.

At 15°C, O2 consumption was identical for the two species. At every other temperature, Z. morio consumed significantly more O2 than A verrucosus. This may reflect a lower standard metabolic rate for the desert species, which is adapted to an environment with limited resources. Neither species showed any sign of stress at either 25°C or 35°C, in that both emerged from experiments active and coordinated, and none died in the three days between experiments.
The two species diverged further at 40°C, with Z. morio dramatically increasing their O2 consumption (note the log10 scale), while O2 consumption in A. verrucosus increased only slightly. Importantly, all Z. morio were visibly impaired at the end of 40°C experiments, showing slow, uncoordinated movement or no movement at all. Eight of twelve Z. morio were dead within two days. In contrast, none of the A. verrucosus tested at 40°C appeared to be stressed, and all survived indefinitely.
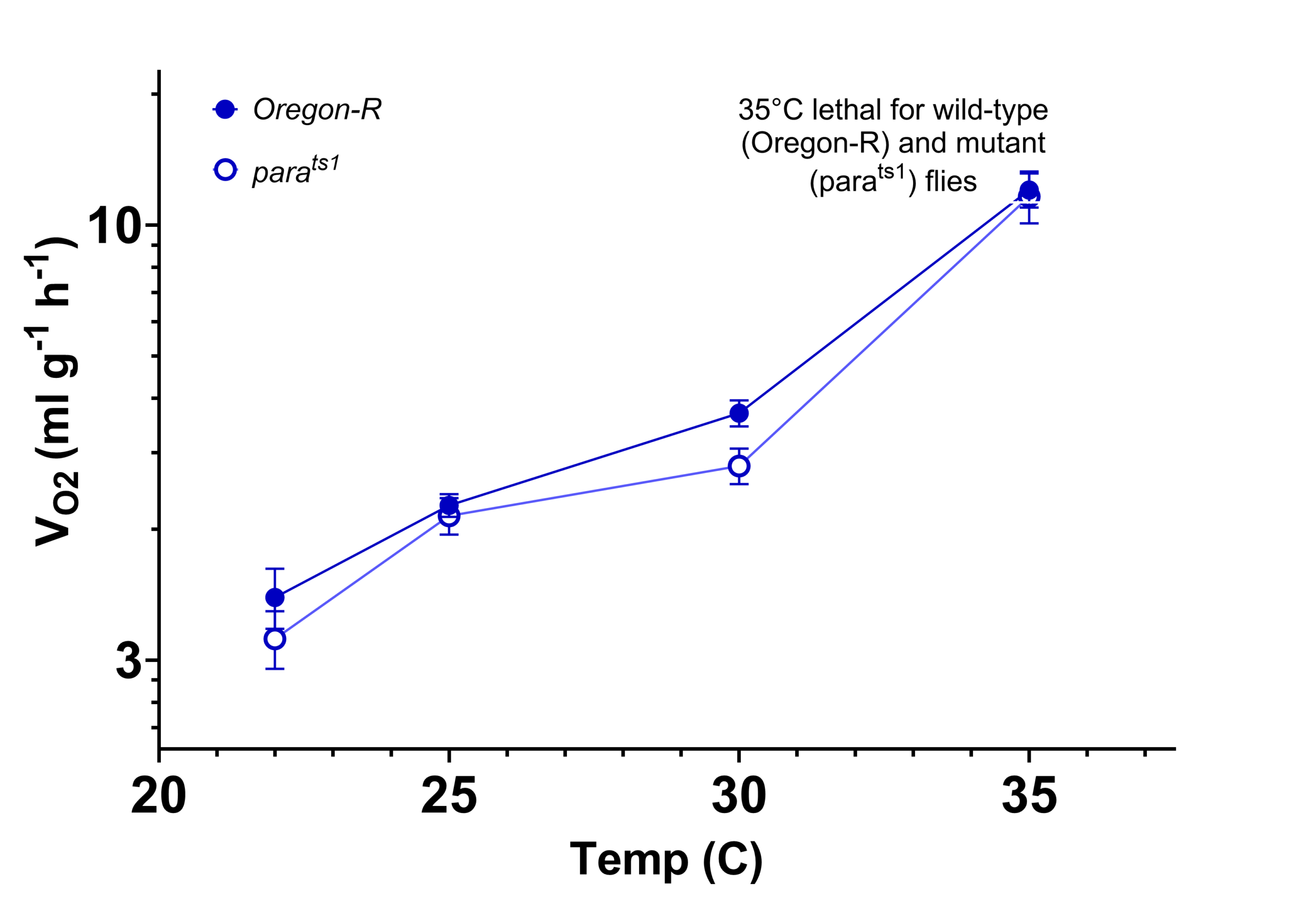
It is perhaps not surprising that Z. morio, a species originating from a relatively constant, resource rich environment, has a higher metabolic rate and is more sensitive to environmental temperature than the desert beetle, A. verrucosus. The dramatic increase in O2 consumption and lethality between 35°C and 40°C for Z. morio was quite striking, however. The lethality-associated increase in metabolic rate resembles that in the fruit fly, Drosphila melanogaster at 35°C (Sandstrom et al., unpublished), and indicates that a dramatic increase in O2 consumption may be a common indicator of severe heat stress in insects.
{kind=link}
There are a few possible caveats. For example, although the two species have been kept under the same conditions for at least four months, which should be enough to reduce the effects of previous physiological adaptations, their life histories as larvae and pupae were different. Z. morio have been reared for multiple generations at the lab, while A. verrucosus were collected from the wild, which may somehow influence their responses to temperature. Asbolus and their relatives can be reared in captivity (Rider, 2024), so it may be worth repeating the experiment with lab-reared beetles.
References
Ahearn, G. A. (1970a). Changes in hemolymph properties accompanying heat death in the desert tenebrionid beetle Centrioptera muricata. Comparative Biochemistry and Physiology 33, 845–857.
Rider, S (2024) Death Feigning Beetles of the United States and Mexico. Publisher S. Rider Jr. 182 pp.